Een organisme is aeroob wanneer het alleen in een zuurstofrijk milieu kan gedijen, omdat het dizuurstof gebruikt ten behoeve van de celademhaling, het energieleverende proces binnen het metabolisme. Strikte aeroben vereisen moleculaire zuurstof (O2) voor de dissimilatie. Hierbij wordt 2880 kJ per mol vrijgemaakt. Deze energie wordt gebruikt om per molecuul glucose 38 ATP-moleculen te regenereren uit 38 ADP-moleculen. Dit is negentienmaal zo veel energie per suikermolecuul als in een typische anaerobe reactie.
Eukaryoten, dat zijn organismen met cellen met een volledige celkern, ontvangen gedurende dit proces slechts een nettowinst van 36 ATP uit het ADP, omdat er energie nodig is om de voedingsstoffen door het celmembraan heen te brengen, via actief transport. Voorbeelden van strikt aerobe bacteriën zijn Serratia en Pseudomonas aeruginosa.
Voor actieve voortbeweging is energie nodig. Deze wordt verkregen door processen zoals de celademhaling of dissimilatie en andere metabolische en enzymatische processen die energiedragende substraten zoals adenosinetrifosfaat (ATP) produceren. Energie wordt opgeslagen in triglyceride in vetweefsel en glycogeen in de spieren en de lever. Metabole spiervermoeidheid kan optreden bij een tekort aan substraten of een teveel aan metabolieten die als afvalstoffen vrijkomen bij de spiercontracties.
Bij langzame duurlopen is er vooral sprake van vetverbranding, bij snellere duurlopen boven de aerobe drempel vooral koolhydraatverbranding. Het energieverbruik per afstand ligt bij de mens bij het rennen bijna tweemaal zo hoog als bij een paard.
Wanneer het zuurstofverzadiging in de mitochondria daalt onder de grens van 60% wordt de anaerobe drempel bereikt. De lactaatconcentratie in de spier stijgt tot 4 mmoles Kg-1 bij het bereiken van de anaerobe drempel.
Bij verhoogde spierarbeid echter, wordt meer pyruvaat verbruikt via de citroenzuurcyclus: hierdoor wordt NAD+ onvoldoende geregenereerd bij de omzetting van pyruvaat naar lactaat.
In een aeroob bloedprofiel moet normaal de lactaat/pyruvaat verhouding kleiner zijn dan 1, de verhouding omega6/omega3 kleiner dan 5, de verhouding Fe/Cu ongeveer gelijk aan 1.
De Mens als Duurloper
Migrerende dieren zoals het paard, de kameel en de gnoe, en jagende dieren zoals de wolf, de hyena en de hond leggen regelmatig lange afstanden al rennend af. Ook de mens is een duurloper en neemt daarmee een bijzondere positie in. Het is de enige primaat die lange afstanden aflegt, en is in vergelijking met andere dieren een slechte sprinter en verbruikt relatief veel energie. Over langere afstanden is de mens echter in staat om de meeste dieren eruit te lopen, hoewel speciaal getrainde paarden en hondenrassen zoals poolhonden meer dan 100 kilometer op een dag kunnen afleggen terwijl ze een mens dragen of voorttrekken.
Als sprinter moet de mens het tegen veel dieren afleggen. Voor langere afstanden geldt dat niet. De mens is niet in staat tot galop, het rennen is meer te vergelijken met draf en daarbij ligt het menselijk tempo aan de hoge kant, zeker als gecorrigeerd wordt voor lichaamsgewicht - hoe hoger het normale gewicht, hoe efficiënter de loop. Hierbij speelt vooral de pasfrequentie een rol die bij kleinere dieren een stuk hoger ligt.
Het hardlooptempo voor langere afstanden varieert bij de mens tussen de 8 en 23 km/u, met de meeste mensen tussen de 11 en 15 km/u. Dit ligt een stuk hoger dan op basis van bovenstaande verhouding voorspeld wordt voor een viervoetige van gelijk gewicht, waarbij het bereik ongeveer tussen de 5 en 14 km/u ligt, met een voorkeurssnelheid van rond de 10 km/u. De mens is echter ook sneller dan grotere dieren, zoals de pony. Voor pony's van 110-170 kg ligt het drafbereik tussen de 7 en 16 km/h, met een voorkeurssnelheid van rond de 11 km/u.
In galop winnen viervoetigen het over het algemeen van de mens. Een viervoetige van 65 kg zou 10-15 minuten rond de 28 km/h moeten kunnen galopperen, terwijl racepaarden de 32 km/u halen over 10 kilometer.
Bij de mens is er wel een verschil tussen het energieverbruik per afstand bij lopen en rennen. Bij de optimale snelheid verbruikt een lopend mens aanmerkelijk minder energie per afstand dan een rennend mens.
De sagittale buiging bij zoogdieren in draaiende galop helpt bij de ademhaling, dit in contrast met de laterale buiging bij hagedissen. Met de benen ver uit elkaar en een holle rug wordt de lucht aangezogen, terwijl de lucht uit de longen wordt gepompt bij de kromme rug als de benen onder het lichaam komen. Een nadeel hiervan is echter dat de ademhaling niet sneller kan dan het looptempo, wat een limiet stelt aan de zuurstofhoeveelheid en de warmteafvoer.
Rennen kan zeer veel warmte opwekken, tot tienmaal zoveel als bij lopen. Dit kan afgevoerd worden door de niet-verdampende koelingsmethodes convectie (stroming), conductie (geleiding) en straling (radiatie). Als de luchttemperatuur echter hoger ligt dan de lichaamstemperatuur, zorgen conductie en straling echter juist voor een verhoging van de lichaamstemperatuur.
Het rete mirabile in de halsslagader werkt volgens het tegenstroomprincipe als warmtewisselaar voor de hersenen en maakt het mogelijk om daar nog warmte af te voeren. Verdampingskoeling werkt ook nog als de omgevingstemperatuur hoger ligt dan de lichaamstemperatuur en de meeste zoogdieren maken hier gebruik van. Daarbij kan weer onderscheid gemaakt worden tussen enerzijds hijgen, waarbij warmte wordt afgevoerd via het ademhalingsoppervlak - neusslijmvlies, mondslijmvlies en tong - en anderzijds zweten, waarbij warmte via het gehele lichaamsoppervlak wordt afgevoerd.
De meeste middelgrote tot grote zoogdieren gebruiken hijgen om warmte af te voeren. Uitzonderingen zijn Hominidae, paarden en kamelen, terwijl zweten bij enkele holhoornigen en kangoeroes ook bijdraagt aan de thermoregulatie. De zweetklieren van varkens en enkele roofdieren lijken geen rol te spelen in de thermoregulatie. De huid van zwetende dieren is koeler en daardoor is de warmteoverdracht groter. Zweten is onafhankelijk van de ademhalingscyclus. Waar de ademhalingscyclus afhangt van de inspanning, is de noodzaak tot warmte-afvoer ook afhankelijk van de omgevingstemperatuur en de vochtigheid.
Lopen wordt benaderd als een omgekeerde slinger waarbij de interactie tussen potentiële gravitatie-energie en kinetische energie het energieverbruik vermindert. In zijn meest eenvoudige vorm is er geen voet en buigen de benen niet, gelijkend op steltlopen. Dit zou een zeer inefficiënte manier van lopen zijn door de abrupte richtingsveranderingen van de snelheid.
Bij rennen wordt tot 70% van de kinetische energie teruggewonnen via elastisch weefsel - collageenrijke pezen en gewrichtsbanden - in de benen. Voor viervoetigen geldt dat bij modellen waarbij de elasticiteit van de rug buiten beschouwing wordt gelaten, de efficiëntie van de galop lager is dan andere gangen, ook bij hogere snelheid. De lange rugspier longissimus thoracis speelt echter een belangrijke rol bij de galop, waarbij de aponeurosis van deze spier elastische energie opslaat, zodat de wervelkolom als een veersysteem werkt. Aangezien de benen sneller naar achteren bewegen dan naar voren, kan dit alleen werken doordat de elastische energie van het ene been via de aponeurosis wordt overgedragen aan het andere been. Hierdoor kan een groot deel van de anders verloren gegane inwendige kinetische energie (IKE) worden teruggewonnen.
De mens verbruikt tijdens het rennen zo'n tweemaal meer energie per massaeenheid dan een zoogdier van vergelijkbare grootte, maar is desondanks een zeer goede duurloper. De skeletspieren en pezen slaan elastische energie op die tijdens de volgende pas deels weer gebruikt wordt. Hierin wijkt de mens echter niet af van andere dieren, zodat op basis daarvan tijdens het rennen een optimale snelheid verwacht zou worden. Wat wel verschilt met andere zoogdieren is de ademhaling. Daarnaast onderscheidt de menselijke huid zich van andere zoogdieren doordat deze grotendeels vachtloos is en meer zweetklieren kent per oppervlakte dan welke andere soort dan ook. Als omnivoor is de mens daarnaast in staat tot koolhydraatstapelen, dit in tegenstelling tot strikt carnivore roofdieren.
Tijdens de evolutie van de mens heeft deze eerst tweevoetigheid ontwikkeld, de eerste stap in het proces van hominisatie, nog voor de ontwikkeling van grotere hersenen. Dit ging gepaard met de nodige Anatomische aanpassingen ten opzichte van de laatste gemeenschappelijke voorouder van mens en chimpansee en de Afrikaanse Hominidae. Bij latere Homo als Homo ergaster, Homo erectus, Homo antecessor en Homo neanderthalensis is er echter duidelijk sprake van strikte bipedie. Daarmee lijkt de strikte tweevoetigheid ergens tussen de 2,5 en 1,8 miljoen jaar geleden tot stand te zijn gekomen. Bij de oudere Homo habilis zijn nog aapachtige kenmerken waar te nemen in de handen. De voeten hebben een voetgewelf en zijn zeer geschikt om tweevoetig op te lopen, maar de enkel suggereert nog een minder gunstige gewichtsoverdracht. Ook het kuitbeen en het scheenbeen zijn al zeer mensachtig. Niet alle eigenschappen zijn even goed terug te vinden bij fossielen. Zo laat de achillespees nauwelijks sporen na, wat het moeilijk maakt deze ontwikkeling te plaatsen. En deze pees speelt een zeer belangrijke rol bij het terugwinnen van elastische energie bij het rennen.
Paslengte is een andere belangrijke factor en anders dan viervoetigen versnelt de mens bij duurlopen vooral door deze toe te laten nemen, pas bij sprinten neemt de pasfrequentie sneller toe. Deze paslengte is veelal meer dan 2 meter en bij zeer goede lopers zelfs meer dan 3,5 meter, ongeveer een meter meer dan vergelijkbare viervoetigen en chimpansees. Bij Homo erectus was de beenlengte mogelijk zo'n 50% groter dan bij Australopithecus afarensis. Hoe langer echter de benen, hoe meer energie het kost om deze te bewegen. Daarbij telt vooral de verder weg gelegen distale massa, zodat kortere tenen bijdroegen aan energiebesparingen. Waar de voet van de chimpansee 14% uitmaakt van de totale massa van het been, is dat bij de mens 9%. De benen wegen bij de mens echter 30% van het totale lichaamsgewicht, terwijl dit bij chimpansees 18% is. Een relatief lage pasfrequentie zal daarom energie besparen.
Bij rennen is de grondreactiekracht - de normaalkracht met daarbij de dynamische krachten - zo'n tweemaal hoger dan bij lopen en kan oplopen tot drie- tot viermaal het lichaamsgewicht. Een deel hiervan wordt opgevangen door het meeveren van het been en de voet, maar het skelet moet de resterende krachten verwerken. Hiertoe zijn de gewrichtsoppervlaktes van de onderste ledematen vergroot. Ook de diafysale belasting - de kracht op het middendeel van een pijpbeen - is groter bij rennen en mogelijk is de dijbeenhals korter om hier het buigmoment te verminderen.
Tweevoetigheid is relatief onstabiel en rennen versterkt dat. Om de stabiliteit van hoofd en romp te vergroten, zijn een aantal spieren en de aanhechtingsplaatsen voor de originis van die spieren vergroot. Vooral musculus gluteus maximus speelt daarbij een belangrijke rol en wordt vooral gebruikt tijdens het rennen. Ook het onafhankelijk kunnen draaien van de romp ten opzichte van hoof en bekken helpt stabilisatie bij het rennen. Tijdens de zweeffase kan er geen gebruik gemaakt worden van grondkrachten die vanuit de heup weerstand worden geboden en leveren de romp met brede schouders en de armen de tegengestelde krachten. Mogelijk heeft de ontkoppeling tussen hoofd en schoudergordel ook voordelen bij het gooien. De korte onderarmen - die ten opzichte van het lichaamsgewicht 50% lichter zijn dan die van chimpansees - leveren hier ook een gewichts- en daarmee energiebesparing op. Om de krachten op het hoofd tijdens het rennen te verminderen, is deze kleiner. Een nekband die niet bij chimpansees voorkomt, maar wel bij rennende dieren als paarden, honden en hazen helpt hier ook bij.
Renjacht is een jachtmethode waarbij prooidieren net zo lang worden achtervolgd door een roofdier totdat ze door uitputting of oververhitting te overmeesteren zijn. De mens kiest voor de renjacht veelal het heetste moment van de dag als deze maximaal profiteert van het ten opzichte van de prooidieren grote vermogen om warmte af te voeren. Door de prooi te achtervolgen met een snelheid die ligt tussen de voorkeurssnelheden van twee gangen kan het prooidier niet optimaal presteren.
Canitrailen vs. Canicross: Een Vergelijking van Aerobe en Anaerobe Inspanning
Regelmatig probeer ik aan geïnteresseerden uit te leggen dat Canitrailen off-road hardlopen met je hond door de natuur is net als Canicrossen, waarbij je in beide sporten gebruik maakt van een heupgordel, een verende lijn, en een trekharnas voor je hond. Canitrailen is geen wedstrijd; het gaat dus niet om snelheid, een snelle tijd of de beste zijn. Canitrailen is meer een soort van uithoudingsvermogen-test waarbij het duurvermogen en de algehele fitheid van jou en je hond worden gemeten. Gedurende de gehele afstand die jullie samen afleggen zijn jullie beiden fit en in goede conditie; jullie mogen geen tekenen van oververhitting, overbelasting of oververmoeidheid vertonen. Door vanuit strategisch oogpunt op tijd te drinken, eten, wandelen of korte pauzes in te lassen voorkom je dat jullie fysiek uitgeput, oververhit of overbelast raken. Tijdens het Canitrailen lopen jullie als team de gehele afstand op een Endurance-tempo die jullie allebei uren achter elkaar moeten kunnen volhouden waarbij er nauwelijks sprake mag zijn van verval.
Een lange afstand lopen zoals een Marathon vereist een andere manier van trainen, ander materiaal en een andere strategie als een Sprint.
Nee. Je hond zit aangelijnd aan je vast omdat jullie door gebieden lopen waar ook veel wild aanwezig is. Wij, Canitrailers, zijn te gast in het natuurgebied en vanuit respect voor de natuur houd je je hond altijd aangelijnd. Als jullie aan elkaar vast zitten zullen jullie met elkaar rekening moeten houden. Vaak is het zo dat de hond in een draf sneller kan als de mens, dus zal de hond als jullie samen hardlopen vaak ‘onder z’n eigen snelheid’ moeten lopen. Dat is voor een hond veel vermoeiender en belastender als het gewoon los met je meelopen. Om lang met elkaar te kunnen canitrailen is het belangrijk om onderweg goed voor je hond en jezelf te zorgen. Vermoeidheidssignalen neem ik altijd serieus en ik geef de hond voldoende gelegenheid om te kunnen drinken, rusten en/of eten; het lopen van een ‘snelle tijd’ op een bepaalde afstand mag nooit ten koste gaan van de gezondheid en grenzen van je hond. Daarom heb ik altijd een trailvest om met in ieder geval een opvouwbare drinkbak voor de hond en een soft flask van minimaal 500ml of bladder van 1,5 liter waarin je water kunt meenemen voor mens en hond.
Het klopt dat zowel het canitrailen als het canicrossen in essentie het samen off-road hardlopen met je hond in de natuur is. Maar daar houdt de vergelijking op. Het canicrossen is explosief, competitief, gericht op sprinten en snelheid waarbij je in het anaerobe energiesysteem loopt. De afstand varieert van een korte cross van 3km tot een lange cross van 5km. Het canitrailen is meer gericht op endurance, uithoudingsvermogen, lange afstanden, energiezuinig lopen, verzorgen en zwerven waarbij je met name in het aerobe energie systeem loopt. Aerobe systeem = Canitrail; Lopen in het aerobe systeem doe je met zoveel mogelijk zuurstof door je uithoudingsvermogen op te bouwen en het hart en longen te versterken (onder je aerobe drempel= omslagpunt waarna je spieren gaan verzuren). Je gebruikt ongeveer 50 tot 80% van je maximale hartslag (zone 1, zone 2 en zone 3) waarbij je ademhaling varieert van gemakkelijk tot licht intens. Anaerobe systeem = Canicross; Lopen in het anaerobe systeem doe je op hoge intensiteit en in explosieve korte stukken waarbij je zuurstoftekort opbouwt zoals in zone 4 en zone 5 oftewel ongeveer 80-90% van je maximale hartslag. De spieren beginnen glucose af te breken. Naast water en kooldioxide geeft het lichaam ook melkzuur als afvalproduct af, waardoor de spieren aanvoelen alsof ze in brand staan.
Honden en hun baasjes racen door het bos tijdens NK Canicross
De Aerobe Capaciteit van Honden
Honden blijken skeletspieren te hebben met superieure oxidatieve capaciteiten. Zoals gezegd hebben honden van nature al meer aerobe spiervezels als mensen. Ik zie dan ook vaak dat mensen meer moeite moeten doen om hun aerobe energiesysteem te trainen als honden. Honden die aan hun baasje vastzitten kunnen meestal niet op hun eigen duurtempo lopen omdat dat voor veel baasjes (nog) te hard gaat. Als je dus met een hond loopt die sneller gaat als jouw basissnelheid onder jouw aerobe drempel, dan zou jij in theorie boven je endurance-vermogen lopen om je hond bij te kunnen houden. Oftewel als je hond in een rustige draf loopt zul jij misschien wel keihard moet sprinten, omdat jullie basistempo's nog te ver uit elkaar liggen. Daarom raad ik het baasje aan om naast het trainen met je hond ook aan de eigen conditie, belastbaarheid en tempo te werken zonder de hond.
Stel je loopt zelf ongeveer 9km/u in Zone 3. Je hond loopt ongeveer 11km/u in een ontspannen draf in zijn Zone 3. Zou je zelf 11km/u lopen dan houd je dat nauwelijks vol zonder te verzuren (Zone 4/5). Maar door technisch ontspannen in die draf van je hond je mee te bewegen, kun jij zelf ook 11km/u lopen met een hartslag en energieverbruik die gelijk staat aan die 9km/u.
Je hebt te maken met hoogtemeters, mul zand, gladde boomwortels, wilde dieren, verschillende weersomstandigheden waardoor je nooit 10km, 20km of 50km continu in hetzelfde tempo kunt blijven lopen. Op de Canitrails is er dus altijd sprake van een grote variatie in externe omstandigheden waarop jullie het tempo continu moeten aanpassen.
Als hardlooptrainer voor honden en mensen verdiep ik mij graag in de basis van het hardlopen. Want waar haal ik en mijn hond de energie vandaan om langere afstanden af te kunnen leggen. Hoe train ik dat dan? Maar nog essentiëler vind ik de vraag of de hond en de mens, beide zoogdieren, dezelfde energiesystemen gebruiken….Mijn zoektocht tot nu toe leverde onderstaande bevindingen op die ik graag met jullie wil delen. Het menselijk lichaam heeft een motor die bestaat uit de spieren en het hart-longsysteem. Als je intensiever gaat inspannen hebben je spieren meer zuurstof nodig omdat er sneller energie vrij moet komen. Daarom gaat je hartslag omhoog bij fysieke inspanning: je hart pompt meer zuurstof naar je spieren. In het begin ga je dieper ademen, maar niet sneller. Er komt vanzelf een punt (als je blijft versnellen) dat je niet meer dieper kunt ademen, maar je bovenbenen wel meer zuurstof vragen. Dat is het moment dat je sneller gaat ademen. Dit is het moment dat de trainingsprikkel begint, boven deze inspanning ga je conditie opbouwen: je aerobe drempel. Dit heet de aerobe drempel omdat je met veel zuurstof vooral je vetten als brandstof aan kunt spreken. Deze intensiteit kun je wel vier tot zes uur volhouden. Blijf je na de aerobe drempel versnellen, dan kom je op je anaerobe drempel.
Om te lopen heeft je motor dus energie nodig. Er zijn vier brandstoffen, namelijk ATP, Creatine Fosfaat, Glycogeen en Vet. Voor de duursporten zijn vooral glycogeen en vet interessant. Het nadeel van vet is dat het langzaam vrij komt en er veel zuurstof nodig is om het vet te kunnen verbranden. Dus vet als brandstof geeft nooit direct energie. De eigenschap van vetten is dat het energiezuinig is. Je doet er lang mee en de verbranding verloopt rustig. Glycogeen, oftewel energiesnelle suikers, vallen onder de tragere koolhydraten. Dit glycogeen ligt opgeslagen in de lever en rond de spieren. Een goed getrainde loper kan op maximaal glycogeen gebruik ongeveer 1,5 uur tot 2 uur lopen. Beide systemen blijven ten alle tijde actief, dus het is een fabeltje dat je ene systeem ( glycogeen) wordt ingeschakeld als je andere systeem (vetten) niet meer meedoet.
In onze spiercellen, om precies te zijn in de mitochondriën, wordt de energie geproduceerd. 1. Het Adenosinetrifosfaat (CP-ATP) systeem is een energiebron voor de hele korte sprint waarbij de energieborn ATP energie en ADP levert. 2. Het Anaerobe Glycolyse systeem is een belangrijke energiebron voor de middenafstanden waarbij het glycogeen wordt afgebroken en daarbij melkzuur en energie levert. 3. Het Aerobe Glycolyse systeem is de voornaamste energiebron voor de lange afstanden waarbij glycogeen met zuurstof samen een enorme energie met koolstofdioxide en zuurstof oplevert. 4. Het Aerobe Vetzuren systeem is de voornaamste energiebron voor de echte diesels waarbij vetzuur met grote hoeveelheden zuurstof wordt afgebroken tot grote hoeveelheden energie, koolstofdioxide en zuurstof. Sprinters gebruiken vooral ATP als brandstof en langeafstandlopers gebruiken de aerobe omzetting van vetzuren en glycogeen.
De inzet van de 4 energiesystemen is de verklaring voor de afname van het vermogen met de tijd die Pete Riegel gevonden heeft. Hoe zit het bij honden? Honden zijn van nature uithoudingsvermogen-dieren. Ze hebben een hoog oxidatievermogen en zijn goed aangepast aan uithoudingsactiviteiten. Er is zelfs een discussie of honden spiervezels hebben die volledig anaëroob zijn! Dat betekent echter niet dat het cardiovasculaire uithoudingsvermogen niet kan worden verbeterd. Om meer te weten over het aerobe of anaerobe vermogen van de hond wil ik verwijzen naar het artikel “Exercise physiology of the Canine Athlete” van Ralp Millard uit 2016. Een goed algemeen begrip over de fysiologische functie van het lichaam van de hond is belangrijk als basis om te begrijpen hoe je hond z'n systemen inzet tijdens het canitrailen.
Honden blijken skeletspieren te hebben met superieure oxidatieve capaciteiten. Het lijkt er volgens onderzoeken dus op dat honden veel minder als mensen gebruik maken van het op glycogeen gebaseerde systeem en meer op het op vetzuren gebaseerde energiesysteem. In plaats van glycogeen gebruiken sledehonden andere energiebronnen zoals spiertriglyceriden (de reeds aanwezige vetvoorraad in de cellen). Honden zijn waarschijnlijk in staat tot deze vorm van metabolisme vanwege hun aangeboren vermogen om vrije vetzuren te gebruiken, hun vermogen om hoge niveaus van training in uithoudingsactiviteiten uit te voeren en de consumptie van een dieet met een hoog percentage vet als een van de componenten. De skeletspier van honden bestaat voornamelijk uit spiervezels van type I, IIA en IIAX, die allemaal in staat zijn tot een hoog oxidatief metabolisme. Zeer aerobe zoogdieren zoals honden hebben een verhoogd vermogen om op vet gebaseerde substraten te gebruiken voor het energiemetabolisme. Het is vrij algemeen aanvaard dat de skeletspier van honden een relatief hoog vermogen heeft tot aëroob metabolisme. Het is waarschijnlijk dat honden veranderingen in het spiervezeltype ervaren als reactie op inspanning. Echter het gebrek aan verandering in capillaire dichtheid in skeletspieren na duurtraining suggereert dat microcirculatie van ongetrainde jachthonden al voldoende is om te voldoen aan de eisen van duurtraining of dat de overbelasting die nodig is om veranderingen te bevorderen in deze onderzoeken niet werd bereikt. Zelfs ongetrainde honden hebben een hoge aerobe capaciteit en lijken genetisch aangepast te zijn voor efficiënte zuurstofextractie en -gebruik. Duurtraining is echter waarschijnlijk zeer gunstig voor de algehele gezondheid van honden en een verbeterde atletische functie.
Krachttraining vergroot de omvang van type II-vezels meer dan type I. Training met hoge weerstand verhoogt het aantal contractiele eiwitten in type II-vezels, waardoor het dwarsdoorsnede-oppervlak en de kracht die ze kunnen genereren, toenemen. Men denkt dat de vergroting van spieren die optreedt bij krachttraining meestal optreedt als gevolg van hypertrofie van spiervezels en niet van hyperplasie (toevoegen van spiervezels). Kracht bij honden heeft voornamelijk te maken met snelheid en het vermogen om lasten te dragen of te trekken. Kracht is ook belangrijk voor honden die zeer snel moeten accelereren en vertragen, zoals honden die deelnemen aan behendigheidswedstrijden. Kortdurende trainingsprogramma’s met maximale intensiteit zijn het meest geschikt om een toename van kracht of snelheid te bevorderen. Naast het conditioneren van de spieren van de ledematen is het ook belangrijk om te focussen op de wervelkolomspieren. Tijdens sprintoefeningen is waargenomen dat de wervelkolomspieren als een van de eersten vermoeid raken. Lopen op een helling op de loopband en bergopwaarts sprinten zijn goede activiteiten om de spieren te versterken, inclusief de wervelkolomspieren. Andere versterkende oefeningen zijn rennen of racen, weerstandsbelastingen trekken of dragen, tegen iets in zwemmen en draven of rennen op hellingen of trappen.
Er is zoals blijkt uit het artikel al veel wel onderzocht maar ook nog heel veel niet. Ik vroeg mij af of de energiesystemen van de mens dezelfde waren als de energiesystemen van de hond. Mijn conclusie tot nu toe is…NEE. Er zijn overeenkomsten maar het energiesysteem van onze lieve viervoeters is anders als die van mensen. Omdat de honden wezenlijk een ander energiesysteem gebruiken tijdens het canitrailen is het natuurlijk ook logisch om de voeding daarop aan te passen.
Spierfysiologie en Aerobe Inspanning
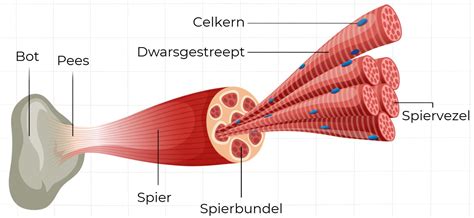
A. Skeletspiercellen spelen een grote rol bij lichaamsbeweging en het bepalen van atletische prestaties, en het is belangrijk om de structuur en functie ervan te begrijpen. Het spierstelsel is een verzameling samentrekkende eenheden die de krachten leveren om verschillende functies uit te voeren, zoals voortbeweging, houding, ademhaling en bloedsomloop. De drie soorten spieren in het lichaam zijn de skeletspier, de gladde spier en de hartspier. Deze sectie richt zich op de skeletspierfysiologie. Skeletspieren worden ingedeeld in typen op basis van de specifieke individuele contractiele en metabolische eigenschappen van elke cel. Een algemeen classificatieschema verdeelt spiervezels op basis van hun contractiesnelheid, wat het resultaat is van hun contractiele en metabolische eigenschappen. Over het algemeen worden spiervezels beschouwd als slow twitch (type I) of fast twitch (type II). Type I-vezels bevatten hogere niveaus van oxidatieve enzymen, myoglobine en mitochondriën en zijn zeer geschikt om aerobe activiteiten uit te voeren en vermoeidheid te weerstaan. Type II-vezels hebben kleinere hoeveelheden oxidatieve enzymen en beperkte aerobe mogelijkheden. Type II-vezels bevatten echter grote hoeveelheden glycolytische enzymen voor het anaërobe metabolisme en zijn in staat hogere spanningsniveaus en verkortingssnelheid te genereren, maar zijn veel minder bestand tegen vermoeidheid dan type I-vezels.
Honden zijn vrij uniek omdat hun snelle vezels, naast de aerobe eigenschappen van slow twitch-vezels, ook aanzienlijke oxidatieve eigenschappen hebben. Er is zelfs significant bewijs dat suggereert dat er geen puur glycolytische spiervezels aanwezig zijn in de skeletspieren van de romp en ledematen van honden. Er is gesuggereerd dat de enige locatie van type IIB-spiervezels bij honden in de larynxspieren (in het strottenhoofd) is. Er wordt aangenomen dat eerdere onderzoeken die alleen op immunokleuring waren gebaseerd, hebben geleid tot een verkeerde classificatie van spiervezels. Nieuwere studies hebben elektroforese, immunoblots, immunohistochemie en beeldanalyse geïmplementeerd om spiervezels van honden te karakteriseren.
Momenteel worden vijf belangrijke vezeltypes beschreven in de romp- en ledematenspieren van honden, waaronder drie zuivere vezels (I, IIA en IIX) en twee hybride vezels (I+IIA en IIAX). De oxidatieve/glycolytische verhouding van hondenspiervezels neemt af van type I (hoogste) naar IIX (laagste). Type IIA is intermediair, met beide hybriden tussen hun respectievelijke zuivere isovormen. De afname van het oxidatieve vermogen van type I naar type IIX is drie tot vier keer minder dan bij andere zoogdieren, wat een andere bevinding is die de superieure oxidatieve capaciteit van de skeletspieren van honden ondersteunt. Zelfs de minst oxidatieve spiervezels bij honden hebben een aanzienlijk vermogen tot aëroob metabolisme in vergelijking met snelle vezels van andere soorten.
B. Vermoeidheid is een disbalans tussen de ATP-behoefte van de spieren en het vermogen om ATP te produceren. Tijdens inspanning neemt de behoefte aan ATP door skeletspieren dramatisch toe. Omdat de componenten die nodig zijn om ATP-niveaus te regenereren uitgeput raken, is er minder ATP beschikbaar om energie te leveren voor aanhoudende spiercontractie, wat resulteert in een onvermogen om te oefenen of te presteren op hetzelfde niveau of tempo als voorheen. Fosfaat is een van de componenten die nodig zijn voor de vorming van ATP. Wanneer ATP-vorming de ATP-consumptie niet kan bijhouden, hopen zich overmatige anorganische fosfaationen op in de spiercel. Hoge concentraties fosfaationen kunnen direct interfereren met de kruisbrugging van actine en myosine, en ook de calciumafgifte uit het sarcoplasmatisch reticulum remmen.
Melkzuur (lactaat) is een eindproduct van het glucosemetabolisme via de glycolytische route. Lactaat wordt gevormd tijdens omstandigheden van onvoldoende zuurstoftoevoer naar spieren of door spiervezels met een laag aantal mitochondriën. Glycogeen is een belangrijke energiebron die tijdens inspanning wordt gebruikt. Uitputting van spierglycogeen is een kritieke factor die verband houdt met vermoeidheid bij topsporters tijdens langdurige inspanning. Sledehonden zijn een goed voorbeeld van een topsporter. Canne-atleten met een hoog uithoudingsvermogen kunnen een opmerkelijk vermogen hebben om de spierglycogeenconcentraties te handhaven tijdens perioden van langdurige inspanning. Op de eerste trainingsdag was er een afname van het spierglycogeengehalte; elk van de volgende trainingsperioden resulteerde echter in een stabiel spierglycogeengehalte, dat hoger was dan het gehalte na de eerste dag. Er is vastgesteld dat spierglycogeenbehoud bij duursporters, zoals sledehonden, optreedt als gevolg van verzwakking van uitputting in plaats van snelle aanvulling. Tijdens het eerste deel van meerdaagse duurraces gebruiken sledehonden een aanzienlijk deel van de spiermassa glycogeen. Op de daaropvolgende dagen gebruiken ze slechts een klein percentage van hun resterende spierglycogeenvoorraden. Honden zijn waarschijnlijk in staat tot deze vorm van metabolisme vanwege hun aangeboren vermogen om vrije vetzuren te gebruiken, hun vermogen om hoge niveaus van training in uithoudingsactiviteiten uit te voeren en de consumptie van een dieet met een hoog percentage vet als een van de componenten.
C. Conditionering van skeletspieren omvat het uitvoeren van fysieke oefeningen die de spieren voorbereiden op de specifieke taak die ze moeten uitvoeren. Training heeft betrekking op het incorporeren van de specifieke oefening in het type sportevenement of activiteit waar de hond bij betrokken zal zijn. Een deel van de training omvat ook gedragsverandering die nodig is om de specifieke activiteit uit te voeren. Tijdens inspanningstraining worden atleten vaak gepusht om een activiteit uit te voeren totdat ze fysiek niet meer in staat zijn om door te gaan. Dit wordt gewoonlijk het overbelastingsprincipe genoemd, dat verwijst naar de noodzaak dat een systeem wordt uitgeoefend op een niveau dat verder gaat dan het effect dat training gewend is. Door het systeem in een overbelaste toestand te plaatsen, past het zich aan nieuwe omstandigheden aan, waardoor het systeem beter in staat is om een taak op een hoger prestatieniveau uit te voeren. Dit principe is van toepassing op het cardiovasculaire systeem, het bewegingsapparaat en andere lichaamssystemen. Factoren die overbelasting beïnvloeden, zijn onder meer de intensiteit, duur en frequentie van een bepaalde oefening. De meeste soorten oefentraining kunnen worden onderverdeeld in duur- of krachttraining.